FAG Originalartikkel
Cellebasert metode for måling av lipidperoksidasjon og antioksidantaktivitet
Cellebasert metode for måling av lipidperoksidasjon og antioksidantaktivitet.pdf(361 KB)
Introduksjon
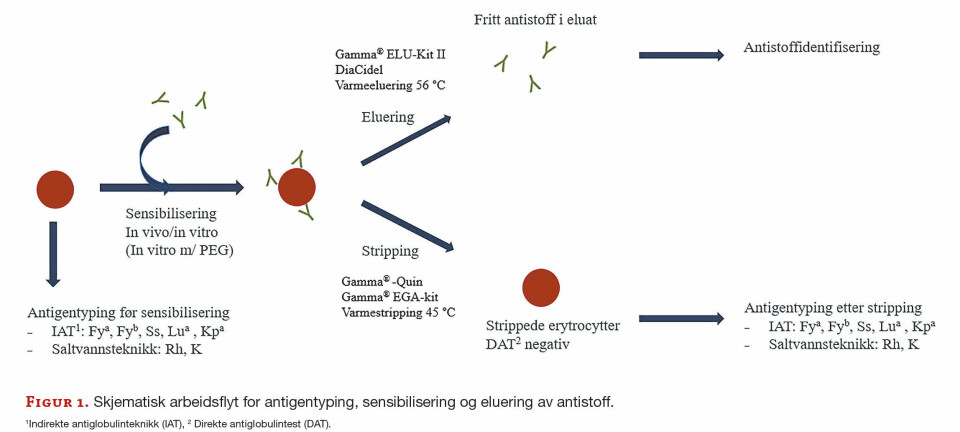
Cellemembranene utsettes kontinuerlig for angrep fra frie radikaler som forårsaker lipidperoksidasjon (Figur 1).
Figur 1
Figur 1: Forenklet modell for lipidperoksidasjon i cellemembraner. Initiering kan skje ved at et fritt radikal X• (HO•, RO•, ROO•, med flere) stjeler et hydrogenatom fra en umettet fettsyre. Reaksjon med oksygen igangsetter en termodynamisk fordelaktig kjedereaksjon med dannelse av reaktive stoffer (sekundære oksidasjonsprodukter) som malondialdehyd (MDA) og 4-hydroxynonenal (4-HNE). Disse kan reagere med biologiske makromolekyler som nukleinsyrer og proteiner, og danne addukter som bl.a. kan forårsake mutasjoner i DNA. Nærvær av katalytiske transisjonsmetaller (redoxsyklende jern- og kopperioner) kan drastisk øke lipidperoksidasjonshastigheten, noe som kan ha betydning da jern akkumuleres i menneskelige organer med økt alder (18).

Lipidperoksidasjonen er forhøyet i degenerative sykdommer som Alzheimers, Parkinsons og multippel sklerose, i sykdommer som involverer jernoverskudd, og ved kronisk inflammasjon (1, 2). I tillegg kan eksponering for sollys skade hudcellenes lysosomer og frigjøre proteinbundet jern, som igjen kan indusere lipidperoksidasjon (3). Ved lipidperoksidasjon dannes reaktive produkter (Figur 1) som kan gi uønskede strukturelle endringer på molekylnivå og være mutagene, det vil si forandre DNA. Slike reaksjoner kan bidra til raskere aldring, celledød og kreftutvikling. Antioksidanter kan imidlertid motvirke slike prosesser (4, 5) og har derfor fått stor oppmerksomhet i mange forskningsprosjekter verden over.
Daglig inntak av frukt og grønnsaker kan redusere forekomsten av sykdommer som aterosklerose, demens, kreft og aldring. Dette kan skyldes at frukt og grønnsaker innholder fytokjemikalier, blant annet potente antioksidanter. Svært få stoffer er imidlertid evaluert, og antioksidanter benyttes ikke i noen større grad i behandling av sykdom. Høye inntak av enkelte antioksidanter (vitamin E, ß-karoten, med flere) kan være skadelig. Antioksidanter finnes i alle organismer og er godt karakterisert i terrestriske organismer, spesielt i planter. Blant annet er mer enn 4 000 polyfenoler identifisert (5). Kunnskapen om antioksidanter i det marine miljø er imidlertid begrenset. Bare i norsk Arktis og subarktiske farvann finnes tusenvis av arter, for eksempel mikroorganismer, virvelløse dyr og planter (alger) som ennå ikke er kjemisk karakterisert. Innenfor marin bioprospektering forsøker vi å finne nye stoffer i marine organismer som kan nyttes til å forebygge og behandle sykdommer.
Identifisering av nye potente antioksidanter krever følsom metodikk som kan håndtere og teste et stort antall prøver av varierende kjemisk natur. Så langt har antioksidantaktivitet hovedsakelig blitt vurdert med kjemiske analyser, som metodene Ferric Reducing Antioxidant Power (FRAP) (6, 7) og Oxygen Radical Absorbance Capacity (ORAC) (8). Analysebetingelsene er langt fra fysiologiske og de tar heller ikke hensyn til om stoffene blir opptatt i cellene. Dette gjør relevansen av resultatene uklare. FRAP gjennomføres for eksempel ved pH 3,6 og omfatter ikke dannelse av frie radikaler. Det er av betydning for bio-prospekteringen at antall falske positive treff i ekstrakter med ukjent sammensetning begrenses.
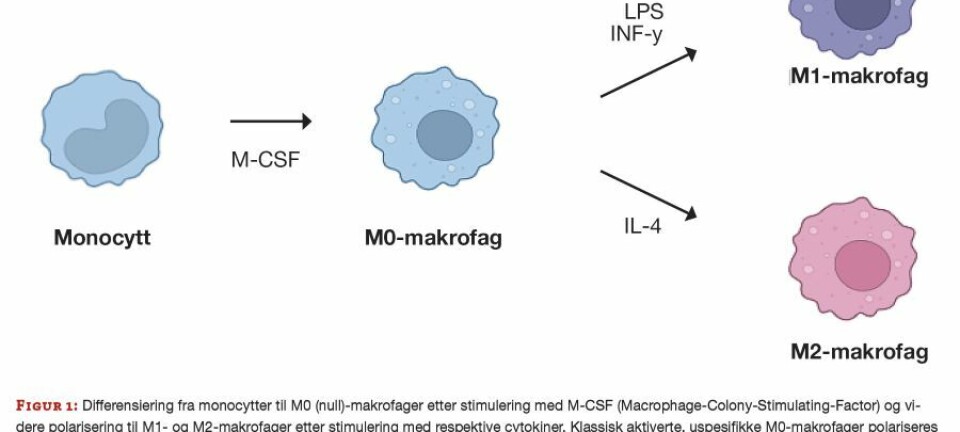
I denne artikkelen beskriver vi en cellebasert mikroplatemetode (Cellular Lipid Peroxidation Antioxidant Activity; CLPAA, vårt akronym) for å måle lipofile antioksidanter og naturlige ekstrakter (skjema 1). Metoden er basert på den cellemembranløselige og oksidasjonsfølsomme lipofile proben C11-BODIPY (9-11) som har lav celletoksisitet (11, 12). Prinsipper for CLPAA-metoden er beskrevet i figur 2.
Skjema

Figur 2
Figur 2: Prinsipper for lipidperoksidasjonsmetoden CLPAA.
A. I CLPAA-metoden tillsettes den lipofile oksidanten cumOOH som i nærvær av redusert jern kan danne hydroksylradikaler (•OH), som vil initiere lipidperoksidasjon (se figur 1) i cellemembraner (9).
B. Den oksidasjonsfølsomme lipofile proben C11-BODIPY er naturlig fluorescerende rød. Ved angrep fra frie radikaler under lipidperoksidasjonen oksideres proben og spaltes ved konjugerte diener (piler), og danner to stabile fluorescerende grønne oksidasjonsprodukter som har mistet en fenylgruppe (19). Fragmenteringen korrelerer til graden av lipidperoksidasjon og måles med fluorescens.
C. Fluorescens-mikroskopi av grønn og rød C11-BODIPY-fluorescens under lipidperoksidasjon i HepG2 celler. Bildene ble tatt cirka én time etter tilsetting av cumOOH og Fe2+. Før eksponering hadde cellene tatt opp 5 μM C11-BODIPY i 30 min. Inkubasjonstemperaturen var 37°C.

For demonstrasjon av metoden sammenlignes jernbinderen desferrioxamine (DFO - en bakteriell siderofor) med flavonoiden luteolin, en polyfenol som finnes naturlig i mange terrestriske planter, inkludert grønnsaker og te. Spesielt te fra gule krysantemumblomster har høy luteolinkonsentrasjon, og teen har interessant nok tradisjonelt vært anvendt som folkemedisin i Asia. DFO er en hydrofil sterk Fe3+-binder (km ~1030,6), men med langsomt celleopptak hovedsakelig ved endocytose. DFO er mye brukt for behandlinger (klinisk kjent som Desferal® som administreres gjennom injeksjon) av jernrelaterte sykdommer, som hemokromatose. På grunn av flavonoidenes hydrofobe karakter kan de binde jernioner, nøytralisere radikaler og passere cellemembraner (13, 14).
Metode
Humane leverceller fra karsinom (HepG2) dyrkes i medium (MEM Earle´s, Biochrom F0325, Berlin, Tyskland) supplert med 10 % fetal bovine serum (FBS), ikke-essensielle aminosyrer, L-glutamin, natriumpyruvat og gentamycin, i 175 cm2 flasker som holdes i vannmettet 5 % CO2-atmosfære ved 37°C. Etter vask av celler med fosfatbufret saltvann (PBS, Mg2+- og Ca2+-fri), trypsineres cellene i 5 min og resuspenderes i cellekulturmediet. Cellekonsentrasjonen bestemmes med telling i Bürkers tellekammer, og om lag 80 000 celler (per 100 μl) blir sådd i svarte 96-brønns polystyrenmikroplater med gjennomsiktig bunn (se skjema 1). Ved å la cellene sette seg på bunnen ved romtemperatur (~30 min) før inkubasjon ved 37°C, minimeres artefakter (15). Etter 24 timer blir eksperimentet utført.
Cellene vaskes med PBS og inkuberes med 5 μM C11-BODIPY (invitrogen D3861, Eugene, OR) i 30 min ved 37°C. Etter skifte til behandlingsmedium (FBS-fri MEM Eagle supplert som ovenfor) tilsettes antioksidanter, og cellene inkuberes ved 37°C i 60 min. Etter en rask PBS-vask tilsettes Hanks saltbuffer (Biochrome L2035), supplert med glukose og 50 μM kumen-hydroperoksid (cumOOH, Sigma-Aldrich) som initierer lipidperoksidasjonen. Grønn og rød fluorescens måles i løpet av en time inne i en plateleser (Wallac VICTOR3, Perkin Elmer) ved 37°C. De brukte lys-filtrene var 485/14 nm (eksitasjons) og 520/10 nm (utslipp) for grønn fluorescens, og 590/7 nm og 632/45 nm for rød. Med innstilling av eksitasjonbølgelengde > 580 nm for rød fluorescens, unngås lysabsorpsjon av C11-BODIPY oksidasjonprodukter som dannes i løpet av eksperimentet (9).
Kontrollbrønner med og uten cumOOH og med kjente antioksidanter (ved screening av fraksjonerte ekstrakter), inkluderes i alle platene. Inkubasjonstider og vaskeprosedyrer kan justeres for å studere effektene av en forbindelse mer detaljert. Antioksidanter kan sammenlignes parallelt ved identiske konsentrasjoner, eller ved å sammenligne IC50-verdier, konsentrasjonen for 50 % hemmende effekt (half maximal inhibitory concentration). Siden antioksidanter kan ha sterkt varierende effekter, gir sammenligning av antioksidanter ved en bestemt konsentrasjon begrenset informasjon. IC50 er et mål på effektiviteten av et stoff, jo lavere verdi jo bedre, og kan bestemmes fra dose-responstester av antioksidanter over store konsentrasjonsintervaller, med tilpassing av sigmoidale dose-responsekurver. Slike kan i tillegg gi informasjon om hvor raskt konsentrasjonsøkningen gir effekt (”Hill slope”). Vanligvis ses en sterk konsentrasjonsavhengighet for potente antioksidanter i μM - mM intervallet (-6 til -3 på en logskala). Selv om full beskyttelse ikke kan oppnås, kan approksimative IC50-verdier fås gjennom å ekstrapolere kurven. Kurveberegninger, hvor bunnplatået settes til y = 0 (som reflekter en perfekt antioksidant), ble utført med programmet Prism 5 (GraphPad Software), ved bruk av sigmoidal kurvetilpasning av data med fire parameters logistisk ligning. Absolutte IC50-verdier er avhengige av celletetthet og andre eksperimentelle forhold. Antioksidanter er følsomme for oksidative modifikasjoner, spesielt ved lave konsentrasjoner. Gjentatte tester av preparater med separate utveiinger og fortynninger bør utføres for å bekrefte resultatene.
C11-BODIPY løses i dimetyl sulfokside (DMSO, Sigma-Aldrich, St Louis, MO) til 6 mM, porsjoneres og lagres frosset for senere tining og umiddelbar fortynning i nevnte behandlingsmedium. DFO (Sigma-Aldrich) og luteolin (Cayman Europe, Tallinn, Estonia) ble løst i vann og etanol og fortynnet i behandlingsmedium. Ved testing av sterkt lipofile prøver og antioksidanter (som carotenoider) blir de som oftest løst i DMSO, og serielt fortynnet i ren DMSO. Til slutt blir 1 μl DMSO tilsatt alle brønner som inneholder 99 μl medium for å gi 1 % DMSO. DMSO og etanol opp til 1 % (v/v) har ikke noen signifikant effekt på lipidperoksidasjonshastigheten, og blir dessuten vasket vekk før tilsetting av cumOOH (skjema 1). CumOOH (88%) oppbevares i kjøleskap og fortynnes umiddelbart for bruk.
Resultater
Fluorescensmikroskopering av C11-BODIPY-inkuberte HepG2 celler viste at lipidperoksidasjonen i fravær av oksidant nesten er ikke-eksisterende (Figur 2C, venstre). Kun rød (og ikke grønn) fluorescens ble påvist etter en times inkubasjon ved 37°C. Tilsetting av oksidanten cumOOH reduserte den røde og økte den grønne fluorescensen noe som tyder på betydelig lipidperoksidasjon. Ved tidligere studier med C11-BODIPY har både nedgangen i rød og økningen i grønn fluorescens blitt brukt til å anslå lipidperoksidasjonen (9, 11). Vi fant at med dyrkede HepG2-celler i 96-brønnsplater, fungerte måling av økningen i grønn fluorescens bedre enn måling av nedgang i rød (Figur 3).
Figur 3
Figur 3: Optimalisering av C11-BODIPY og cumOOH konsentrasjoner for den cellebaserte lipidperoksidasjonsmetoden CLPAA. HepG2-celler (80 000 per brønn, 24 timer etter utsåing) tok opp C11-BODIPY i indikerte konsentrasjoner (0-10 μM) i løpet av 30 min ved 37°C. Etter vask med PBS ble det tilsatt cumOOH og (A) grønn og (B) rød fluorescens ble målt. Gode signaler fra dannelse av grønne C11-BODIPY oksidasjonsprodukter ble målt mellom 3-10 μM C11-BODIPY. CumOOH-konsentrasjon over 50 μM ga bare liten økning i peroksidasjonen. For rød fluorescens viste det seg at plasten på 96-brønnsplatene (Costar 3603) ga sterkt signal allerede uten tilsetting av C11-BODIPY ved samme lysintensitet som for grønn fluorescens (kun resultatene for 0 og 10 μM C11-BODIPY vises). Tilsetting av C11-BODIPY økte som den bør, og bakgrunnssignalet var signifikant. Fra dette eksperimentet ble 5 μM C11-BODIPY og 50 μM cumOOH valgt for rutinemessig test av antioksidanter. Stigningskoeffisienter (indikert med α0 og α50) fra lineær regresjon for økningen i grønn fluorescens i det initiale lineære tidsintervallet 0-40 min brukes som mål på lipidperoksidasjonshastigheten. Data er fra avlesning hvert tredje minutt der punkter motsvarer gjennomsnittverdien av N = 4 avlesninger.

Nedgang i rød fluorescens var relativt lav i forhold til økningen i grønn fluorescens, og høye bakgrunnssignaler fra plasten gir det røde fluorescensområdet lavere følsomhet enn det grønne. Vi undersøkte også om forholdet grønn/rød fluorescens ved endepunktet kunne brukes, men fant ut at visse forbindelser påvirker (absorberer) grønne og røde lyssignaler forskjellig. Dette gir en uønsket ikke-lineær effekt som er spesielt tydelig ved høyere konsentrasjoner.
Tilsetting av jern (Fe2+) økte den cumOOH-induserte lipidperoksidasjonshastigheten i HepG2 cellene (Figur 2C, høyre). Celler dyrket i medium og supplert med serum, har vanligvis tilstrekkelige mengder labilt jern (om lag 5 μM i cytosolen hos hepatocyter (16) - mitokondrier og lysosomer har høyere innhold). Siden tilsetting av katalyserende jern vil kreve høyere konsentrasjoner av antioksidanter for samme beskyttelse og gi mindre naturlige forhold, undersøkte vi om tilsetting av ekstra jern var nødvendig for cumOOH indusert lipidperoksidasjon. Resultatene viste at ekstra tilsatt jern ikke behøvdes ved cumOOH indusert lipidperoksidasjon.
Hvor mye cumOOH og C11-BODIPY man skal bruke, avhenger blant annet av cellekonsentrasjon, inkubasjonstiden og instrumentets følsomhet. Konsentrasjonstester med C11-BODIPY (Figur 3) viste stabil initial lineær økning av grønn fluorescens etter cumOOH tilsetning ved 3-10 μM C11-BODIPY (80 000 HepG2 celler per brønn), men 10 μM ga ingen betydelige fordeler over 5 μM, og ved konsentrasjoner ≤ 1 μM C11-BODIPY blir det, ut fra våre erfaringer, vanskelig å skille de røde og grønne signalene fra bakgrunnen (celler, plast). Konsentrasjonstester med cumOOH viste videre at peroksidasjonshastigheten ikke nødvendigvis er relatert til konsentrasjonen av cumOOH, men i større grad kan være avhengig av innholdet av lipidassosierte, katalytiske metaller, mest sannsynlig assosierte jernioner. Inkubasjon med 5 μM C11 -BODIPY i 30 min for inkubasjon med antioksidanter og eksponering for 50 μM cumOOH ble valgt som standard forhold (skjema 1).
Ved direkte sammenligning mellom DFO og luteolin, ga luteolin sterkest antioksidativ beskyttelse, med fullstendig stopp av lipidperoksidasjonen ved høye konsentrasjoner (Figur 4). Andre kommersielle antioksidanter og ekstrakter (HPLC fraksjonerte) fra marine organismer har blitt testet i CLPAA-metoden med tilfredsstillende reproduserbare resultater.
Figur 4
Figur 4: Redusert lipidperoksidasjonshastighet ved tilsetting av antioksidanter.
A. Kjemiske strukturer for testede stoffer. Når DFO, som er et lineært hydrofilt positivt (+1) ladet molekyl ved pH 7.4, hexadentatebinder Fe3+ erstattes tre protoner og ferrioxamine (FO, +1 ladet ved pH 7.4) dannes (venstre). Luteolin, en hydrofob uladet flavonoid, kan binde fritt Fe2+ eller Fe3+ på forskjellige plasser (høyre) (4, 13), og kan i tillegg nøytralisere frie radikaler gjennom å donere et hydrogenatom (H•) eller et elektron (e-) (13). Blant annet kan et uladet luteolin-Fe(II) kompleks dannes (uladete molekyler passerer cellemembraner lettere).
B. Dose-responstester av DFO og luteolin i CLPAA-metoden. HepG2-celler (80 000 per brønn, 24 timer etter utsåing) tok opp 5 μM C11-BODIPY i 30 min, fulgt av én time inkubasjon med DFO og luteolin. Etter en PBS-vask ble 50 μM cumOOH tilsatt for å indusere lipidperoksidasjon. Dannelseshastigheten av fluorescerende grønne C11-BODIPY oksidasjonsprodukter (y-akse) i det lineære intervallet 0-40 min, ble redusert med økt innhold av antioksidantene (x-aksen er logaritmisk). Hver konsentrasjon (0-400 μM) ble testet i triplikat (N = 6 for kontrollene), og hele eksperimentet kunne utføres i en 96-brønnsplate. Sammenligningen viser at luteolin (IC50 = 5,21 μM) var mye bedre til å forhindre lipidperoksidasjon enn DFO (IC50 = 784 μM).

Diskusjon
Tradisjonelt har lipidperoksidasjon blitt studert i biologiske systemer in vitro ved å bruke preparerte mikrosomer fra rottelever, hvor dannet malondialdehyd (reaktivt oksidasjonsprodukt) reagerer med thiobarbitursyre (17). Denne prosedyren har ofte fått kritikk for å være uspesifikk og ukomplett, og kan vanskelig brukes for studier av lipidperoksidasjon hos celler dyrket i mikroplater. CLPAA-metoden benytter den oksidasjonsfølsomme proben C11-BODIPY, som er godt egnet til studier av oksidasjonstilstanden i intakte membraner hos levende celler dyrket i mikroplateformatet. CLPAA-metoden er beskrevet for leverceller, men kan sannsynligvis brukes også på andre celletyper.
Det bør noteres at antioksidanter kan påvirke fluorescensen gjennom lysabsorpsjon, men ved måling av endring av lipidperoksidasjonshastighet som i CLPAA, trenger man ikke å korrigere for antioksidantens egen lysabsorbans.
Våre celler beskytter seg mot høye frie transisjonsmetallnivåer. Tilsetting av ekstra jern i metoden er ikke nødvendig, noe som innebærer eksperimentelle forhold nærmere de fysiologiske. Vi har funnet at metoden også kan brukes for å påvise stoffer som øker lipidperoksidasjonshastigheten (prooksidanter), men vi viser ikke disse dataene i denne artikkelen.
Oppsummering
En følsom metode for testing av antioksidanter i en cellebasert lipidperoksidasjonsmodell har blitt utviklet. Metoden måler hastigheten for dannelse av fluorescerende grønne C11-BODIPY oksidasjonsprodukter. Ved at hastigheten reduseres ved tilsetning av antioksidanter, kan virkningsgraden (antioksidativ kapasitet) til antioksidanter bestemmes. Denne beskrevne CLPAA-metoden vil være et godt supplement til eksisterende metoder (de kjemiske metodene FRAP og ORAC, samt de cellebaserte CAA og Comet-metodene) vi bruker for å finne ukjente antioksidanter ved vår pågående screening av ekstrakter fra marine organismer. Ved CLPAA-basert screening har vi funnet flere stoffer med høy aktivitet i fraksjonerte marine ekstrakter. Stoffene blir nå opprenset og kjemisk karakterisert, og kan forhåpentligvis komme til nytte.
Takk til
Norges forskningsråd og vår samarbeidspartner ABC Bioscience for finansiell støtte av prosjektet. Fluorescensmikroskopibilder ble tatt ved Universitetet i Tromsøs kjernefasilitet Bioimaging.